Крупные тектонические движения на границе позднего Плейстоцена (джунгарская фаза) наряду с новым увлажнением климата вызвали бурное развитие эрозионно-аккумулятивных процессов. В высокогорных областях увеличивавшиеся ледники вновь поползли по долинам, придавая им форму трогов, расположенных на более низком уровне. Два-три конечноморенных вала большей частью хорошо выражены в устьевых частях неоплейстоценовых трогов. Достаточно четко сохранились и ледниковые кары этой эпохи. На Алтае выделяют две стадии оледенения: Чибитскую и Менскую.
В Сауре оледенение именуют Уластинским. В Джунгарии и Северном Тянь-Шане, где следы этого оледенения встречаются в высокогорьях повсеместно, оно может быть названо соответственно Джунгарским и Тянь-Шаньским. Оледенение здесь также прошло стадии сокращения и расширения, о чем свидетельствуют конечные морены. У подножий горных массивов в размытые среднеплейстоценовые конусы выноса были вложены (ныне прекрасно выраженные морфологически) также валунно-галечные конусы выноса из углублявшихся речных долин, в которых сформировался комплекс первых надпойменных террас.
В это же время на опускавшихся участках верхнеплейстоценовый аллювий налегает на среднеплейстоценовый, что наиболее часто наблюдается в горах Алтая. Неравномерности тектонических поднятий здесь повлекли за собой смещение Иртыша в районе р. Шульбы на 5—7 км к северу от его древней долины, выполненной неоген-среднеплейстоценовыми отложениями, и выработку в скальных палеозойских породах новой долины. За пределами Алтая долина Иртыша в описываемую эпоху приобрела современное направление, окончательно оформилось Обь—Иртышское междуречье. Здесь на территории Казахстана верхнеплейстоценовые аллювиальные отложения слагают в долине Иртыша первую (7—10 м), вторую (15—20 м) и третью (22—26 м) надпойменные террасы. В них нередко встречаются остатки мамонтовой фауны.
В результате тектонических дислокаций джунгарской фазы произошли существенные изменения в бассейне р. Чу. Эта река берет начало в Киргизии, но большей частью протекает по территории Казахстана и влияет на палеогеографическую обстановку, в частности на заполнение Аральской котловины. Поэтому вкратце коснемся изменений, происходивших на территории Киргизии. В течение раннего и среднего плейстоцена р. Чу впадала на западе в оз. пра-Иссык-Куль, образуя мощную валунно-галечную дельту, сохранившуюся в виде останца гор Босбармак (1814 м). На востоке в это время формировалась обширная аллювиально-озерная равнина, ныне прорезанная реками Тюп и Джергалан на глубину до 50—60 м. Из отложений, вскрытых р. Джергалан, собраны остатки слонов, носорогов, оленей, бизонов и других животных, обитавших здесь в раннем и среднем плейстоцене. В эту же эпоху верховьем р. Чу, протекавшей по Казахстану, служила р. Чон-кемин, берущая начало с горного узла, соединяющего хребты Заилийский и Кунгей-Алатау. Последний на западе сливался с Киргизским хребтом, замыкая с севера Кокмойнокскую котловину— узкий залив оз. пра-Иссык-Куль. Когда перемычка между хребтами была тектонически раздроблена, воды пра-Иссык-Куля частично стекли по Буамскому ущелью, уровень озера понизился до современного (1608 м), сократилась его площадь. Чу (верховья) несколько отклонилась к западу и через высохшую Кокмойнокскую котловину и Буамское ущелье потекла к северу в Казахстан, а р. Чонкемин стала ее правым притоком. С этого времени она стала полноводнее.
В это же время окончательно прорезались ущелье Капчагай и древнеплейстоценовый конус выноса р. пра-Чилик, что перегораживал Илийскую впадину на меридиане гор Калкан. Водные потоки, стекавшие в Илийскую впадину с хребтов Тянь-Шаня и Джунгарии, образовали р. Или, за ущ. Капчагай она вышла на Южно-Прибалхашскую равнину. Эта равнина, слегка увлекаемая в поднятие совместно с Джунгарской горной системой, уже полностью освободилась из-под покрывавших ее ранее озерных вод, которые ингрессировали в межсо-почные понижения денудационного рельефа периферии Центрально-Казахстанского щита. При этом некоторые сопки оказались окруженными водами озера и образовали острова, столь характерные для северо-западного побережья — ур. Мынарал и др. Здесь на скалистых обрывистых берегах можно видеть белые карбонатные налеты, фиксирующие наиболее высокое стояние уровня воды озера. Они расположены на 6 м выше современного уровня (342 м). На таком же уровне, с абсолютной отметкой 348 м, расположены на равнинных межсопочных участках древние-позднеплейстоценовые береговые валы. Так в позднем плейстоцене начал формироваться Балхаш с конфигурацией, близкой к современной. Прорезали равнину, устремившись к Балхашу, и реки, стекавшие с гор Джунгарии: Каратал, Биен, Аксу, Лепсы. Во всех речных долинах аллювий формировал первые надпойменные террасы. У северного подножия Заилийского Алатау аллювиально-про-лювиальные отложения алма-атинской свиты сливаются с первой надпойменной террасой р. Или. В тех и других отложениях были найдены остатки мамонтов.
В Алакольской впадине, отделяющей Джунгарскую горную систему от хр. Тарбагатай, оформились в конфигурации, близкие к современным, и вытянутые в северо-западном направлении озера Джаланашколь, Алаколь, Уялы, Сасыколь, Кылы (ныне солончак). От озер к северо-восточному окончанию Балхаша неширокой полосой протягиваются песчаноглинистые аллювиально-озерные отложения верхнеплейстоценовой колдарской свиты, мощностью 5—6 м., Местами в них содержится обильная фауна моллюсков.
В широких древних долинах Центрального Казахстана, разобщенных обширными мелкосопочными и пенепленизиро-ванными междуречьями, первые и вторые надпойменные террасы сложены суглинисто-песчано-щебнистым материалом, нередко плохо отсортированным, явно пролювиального генезиса. Все это свидетельствует, что в позднем плейстоцене реки здесь, как и ныне, нередко имели прерывистое течение, пересыхали, их террасы формировались за счет временных потоков. Только реки, подобные Сарысу, Ишиму, отлагали в своих долинах аллювий, слагающий первую и вторую надпойменные террасы. Из второй террасы Ишима известны многочисленные остатки мамонтовой фауны.
В Тургайском прогибе, как показали исследования А. А. Бобоедовой, в начале ширтинского межледниковья происходил интенсивный сток вод из Западной Сибири к Аральскому морю по сквозной ложбине, образовавшейся еще в эпоху самаровского оледенения. В то время ложбина была выработана на максимальную глубину. Позже, в эпоху казанского межледниковья, когда возродился сток вод сибирских рек в Северный Ледовитый океан и при этом понизился базис эрозии, прекратился сток вод по сквозной ложбине. По южной части Тургайского прогиба, куда стекали водные потоки с окружающих возвышенностей, продолжался сток к Аральской котловине.
В это же время прекратило свое существование Восточно-Кызылкумское озеро, образовалась долина р. Сырдарьи и воды ее устремились также к Аральской котловине. Верхнеплейстоценовые отложения, слагающие в долине Сырдарьи первую и вторую надпойменные террасы, выделяются в голод-ностепскую свиту. Этот аллювий близ Арала состоит преимущественно из мелкозернистых песков буровато-серой окраски с прослоями разнозернисто-гравийных песков и песчанистых глин, местами загипсованных. На юге, у границы Казахстана, преобладают гравийно-галечные накопления, перекрытые палево-серыми макропористыми лёссовидными суглинками. Общая мощность обычно 50—60 м, но в районе впадения р. Арысь она достигает 100 м. В первую половину позднеплейстоценовой эпохи р. Сырдарья принимала справа крупный приток вод слившихся рек Чу и Сарысу. Таким образом, есть основание полагать, что в это время началось заполнение водой Аральской котловины.
В позднем плейстоцене прекратился сток вод Амударьи к Каспию через Низменные Каракумы, прорезалось русло по Западным Кызылкумам и воды ее потекли на север, но, по исследованиям А. С. Кесь, не достигли Аральской котловины. Воды Амударьи, заполнив Хорезмскую и Сарыкамышскую впадины, потекли через Узбой к Каспийскому морю; в Аральскую котловину они стали поступать лишь в голоцене.
На Прикаспийской низменности верхний плейстоцен начинается прибрежно-морскими верхнехазарскими слоями: галечниками, песками, ракушечниками с Didacna surachanica и другими моллюсками. В междуречье Волга—Урал они залегают с размывом на абрадированной поверхности континентальных суглинков. Затем отлагаются пески и суглинки регрессивной серии и начинается широко распространившаяся хвалынская трансгрессия моря, отложения которой подразделяются на нижнехвалынские пески и глины, мощностью 5— 10 м с Didacna ebersini, и верхнехвалынские слои, представленные песками (мощностью до 10 м) с Didacna praetrigonoides и другими моллюсками. Почти повсеместно эти пески позже были переработаны ветром.
Вторая половина позднеплейстоденовой эпохи, как и предыдущих плейстоценовых эпох, отличалась аридностью климата. В это время у подножий горных массивов отлагался мелкоземистый супесчаный или суглинистый материал. Обычно он перекрывает ранее отложившиеся пески и галечники, иногда налегает на различные более древние породы, а в западных отрогах Алтая наращивает среднеплейстоценовый лёссовый плащ; содержит остатки мамонтовой фауны.
Насколько грандиозны были описанные здесь преобразования Казахстана к концу позднеплейстоценовой эпохи хорошо видно при сопоставлении палеогеографических карт среднеплейстоценовой эпохи (см. рис. 21) и позднеплейстоценовой (см. рис. 22 — см. с. 76).
В позднем плейстоцене на территории Советского Союза сложился мамонтовый, или верхнепалеолитический, фаунистический комплекс. Главными представителями его являются мамонт, шерстистый носорог, бык-тур и короткорогий бизон. Продолжали существовать многие животные, появившиеся ранее, некоторые из них закончили свой жизненный путь в эту эпоху.
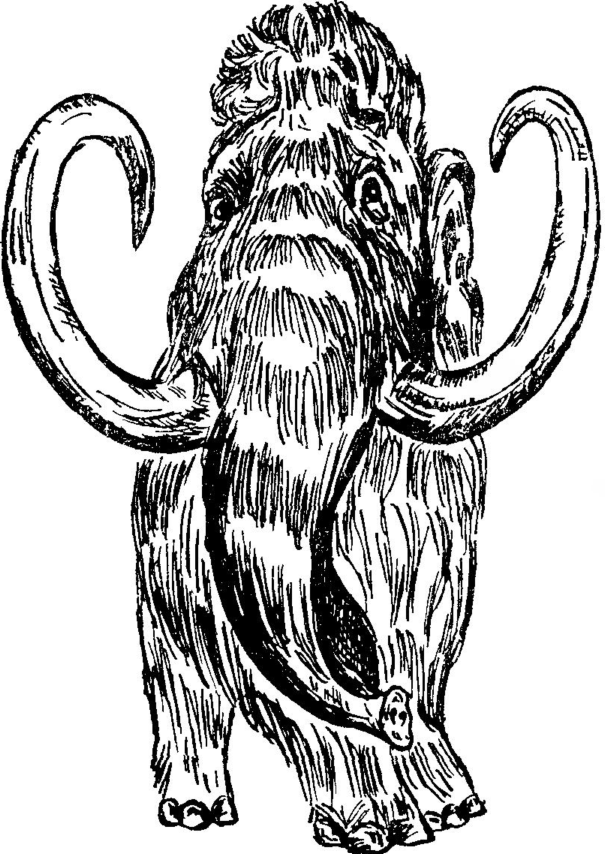
Мамонт Mammuthus primigenius Blumenbach, 1799 (рис. 27) [Mamontheus primigenius; Elephas primigenius; Elephas mamontheus; Elephas jasnovskii]. Этот мамонт — последний представитель хоботных, обитавших в Казахстане. В вечно мерзлой почве Сибири еще до сих пор находят тела мамонтов. Мясо одного из них было настолько свежее, что его могли есть собаки, а в желудке сохранилась непереваренная пища из хвои и ветвей листопадных и лиственных деревьев: ивы, березы, ольхи. Шерсть у мамонтов густая и длинная, бурого цвета, она хорошо предохраняла животное от холодов, как и толстый слой подкожного жира. Шерсть покрывала и уши, которые у мамонта были небольшими, а также — короткий хвост. Хобот тоже был покрыт шерстью. Конец его отличался от конца хобота ныне живущих слонов сильным развитием верхней губы, наподобие ушной мочки.
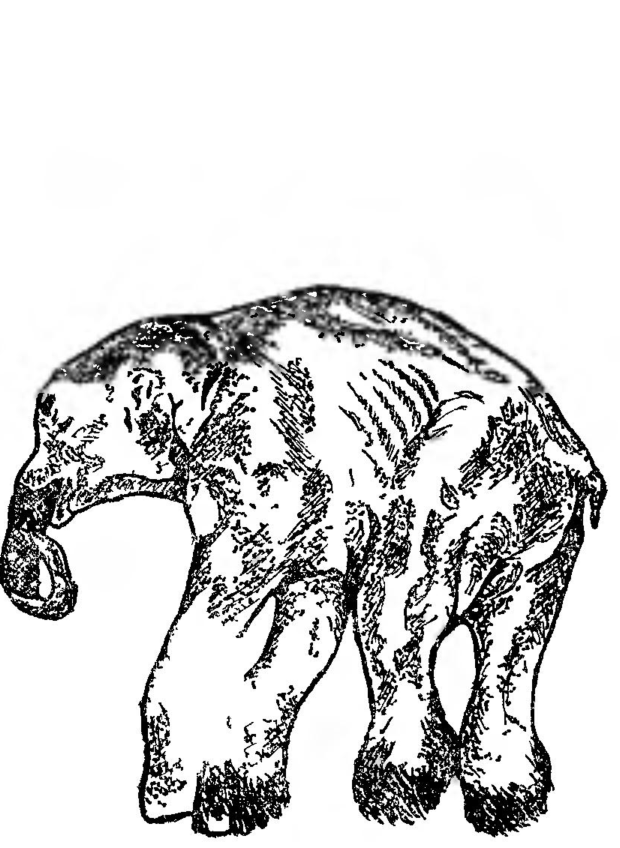
Бивни, достигавшие в длину 3 м, представляли собой два видоизмененных верхних резца, четыре коренных зуба (больших и высоких) хорошо приспособлены для пережевывания грубой растительной пищи. Благодаря сильно развитым бивням, голова у мамонта была крупной, больше, чем у современных слонов. В высоту мамонт достигал 3,5 м, масса — 6 т Недавно в вечной мерзлоте был найден прекрасно сохранившийся мамонтенок, названный Димой (см. рис. 28).
{kind=link}
{kind=link}
Костные остатки мамонта приурочены к верхнеплейстоценовым отложениям. Наиболее часто они встречаются в северной половине Казахстана: в Прииртышье, Приишимье и низовье р. Урала. Самые южные находки известны из аллювиально-пролювиальных отложений, на которых расположен г. Алма-Ата, а также из аллювия р. Джергалан, восточнее юз. Иссык-Куль. Следовательно, мамонты паслись среди горных хребтов Северного Тянь-Шаня.
Шерстистый носорог Coelodonta antiquitatis Blumenbach, 1799 (Rhinoceros tichorhinus Fischer — рис. 29). Этот двурогий носорог, покрытый длинной рыжевато-бурой шерстью, представлял собой приземистое животное на коротких ногах с большим загривком и длинной головой. Длина туловища — до 3,6, высота — до 1,7 м. Передний — носовой рог — у старых особей достигал 1 м длины и массы — до 15 кг.
{kind=link}
Этот рог наряду с копытами использовался животным для добывания корма зимой из-под снега; он был очень мощным. Задний рог — лобный — был менее мощным. Рога сидели на утолщениях носовых и лобных костей и достигали у основания 25 см в поперечнике. Рога были как у самцов, так и у самок. Коренные зубы высококоронковые, с цементом. Резцов не было ни в верхней, ни в нижней челюсти; и траву и ветви, которыми питался носорог, он захватывал ороговевшими краями губ. В зубах носорогов с Вилюя и Индигирки были обнаружены ветки ивы и хвойных деревьев, а спорово-пыльцевые спектры из проб с зубов показали преобладание пыльцы злаков, осок и сложноцветных. Шерстистый носорог придерживался открытых пространств с травянистой и кустарниковой растительностью, перемежающейся с островными лесами.
Костные остатки шерстистого носорога нередко встречаются совместно с остатками мамонта в верхнеплейстоценовых отложениях северной части Казахстана; они известны на Алтае и в Центрально-Казахстанском мелкосопочнике, но на юге пока не обнаружены. Однако кости этого носорога недавно были найдены даже в Южном Таджикистане в фауне палеолитических стоянок Огзи-Кичике (Ранов и др., 1973). Поэтому не исключена возможность обитания шерстистого носорога одновременно с мамонтом и в Южном Казахстане.
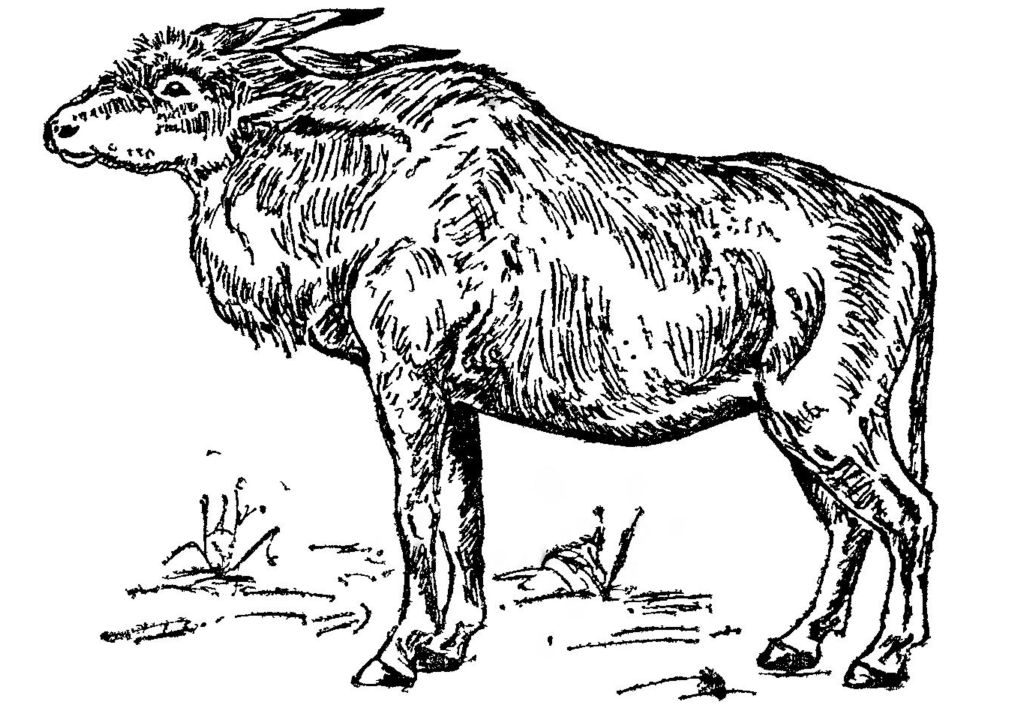
Лошадь Equus caballus sub. sp. Фоссилизированные кости лошадей нередко находят вместе с костями других животных мамонтового фаунистического комплекса во многих районах Казахстана. Несомненно, что позднеплейстоценовые лошади были повсеместно распространены в Казахстане. Это были крупные животные с массивными метаподиями и широкими копытами.
Кабан Sus scrofa Linnaeus, 1758. Зубы дикого кабана найдены археологом X А. Алпысбаевым в пещере Караунгур на правобережье р. Арысь, где жил палеолитический человек.
Косуля Capreolus capreolus, Linnaeus, 1758. Кости косули найдены в пещере Караунгур, а также на Алтае на р. Бухтарме.
Лось Alces alces Linnaeus, 1758. Позднеплейстоценовый лось был очень сходен с современным видом. Фоссилизированные кости лося найдены на Алтае, в Северном Казахстане на р. Чаглинке и на р. Урале.
Северный олень Rangifer tarandus Linnaeus, 1758. Остатки северного оленя в совместном захоронении с остатками мамонта, шерстистого носорога и других животных мамонтового фаунистического комплекса известны из аллювия второй надпойменной террасы Ишима, из аллювия низовий р. Урала. Это типичный вид полуоткрытых болотистых ландшафтов, на что, в частности, указывает строение его копыт. Ареалом ископаемого северного оленя нельзя считать лишь места с очень холодным климатом, так как в прошлом столетии он еще обитал в Казахстане до параллели 50° с. ш.
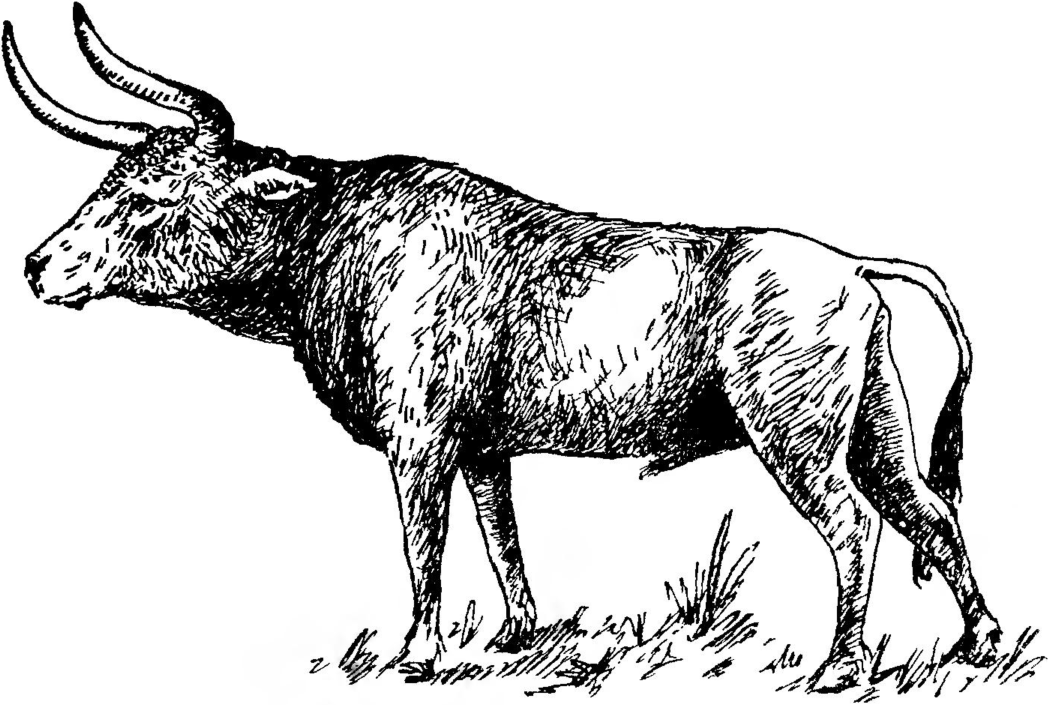
Бык-тур, или первобытный тур, Bos (Bos) primigenius Bojanus, 1827 (рис. 30). Это было крупное животное, значительно превосходившее размерами домашнего быка или зубра.
{kind=link}
Высота его в холке достигала 1,8 м, а масса доходила до 1,5 т. Однако по общему облику он выглядел легче и стройнее наших домашних быков. Рога у тура были большие, длиной до 2 м, очень острые, они служили ему хорошей защитой от врагов. Он был обитателем преимущественно степных увлажненных ландшафтов, хорошей пищей его обеспечивали прибрежные луга. В близких условиях обитал и короткорогий бизон, но он придерживался и лесной зоны.
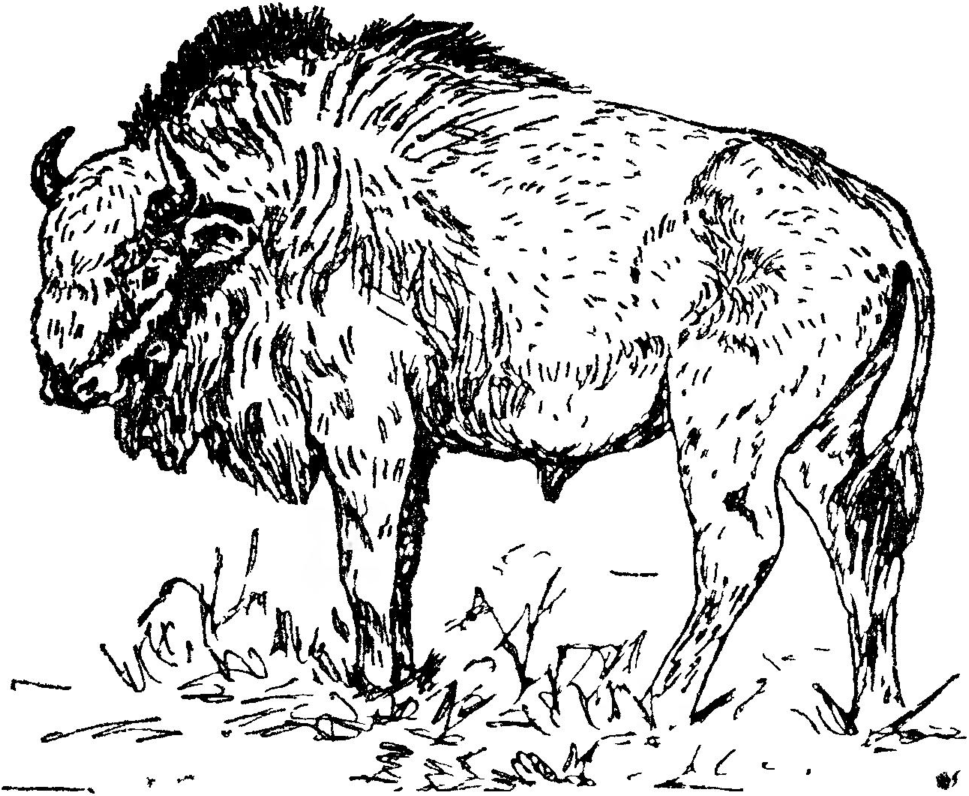
Короткорогий бизон Bison (Bison) priscus mediator Hil-zheimer, 1918 (Bison priscus deminutus V. Gromova, 1935; Bison bonasus L. (рис. 37). У этого бизона череп был значительно короче, чем у длиннорогого. На нем были расположены более толстые и сильно изогнутые роговые стержни. Расстояние между вершинами рогов составляет 67\’—90, а обхват основания — 26—33 см. Характерным отличием описываемого бизона от среднеплейстоценового является также тонкость роговых стержней, меньшая сплющенность их, значительное захождение за лоб с отклонением наверх.
{kind=link}
Костные остатки короткорогого бизона и быка-тура очень часто встречаются совместно с остатками мамонта и шерстистого носорога в верхнеплейстоценовых отложениях, особенно в северной части Казахстана.
Байкальский як Bos (Poephagus) baikalensis N. Verestchagin, 1956. Вероятно, внешне он был похож на современного тибетского яка. Это длинношерстный хрюкающий, а не мычащий бык придерживался каменистых высотных пастбищ. Кости яка обнаружены на Алтае близ устья р. Бухтармы.
Кяхтинский винторог Spirocerus kjaktensis М. Pavlova, 1910 (рис. 32) Эта довольно крупная антилопа, видимо, не способная к быстрому бегу, была обитательницей полуоткрытых горных ландшафтов Алтая в позднем плейстоцене.
{kind=link}
В верхнеплейстоценовых отложениях различных районов Казахстана найдены костные остатки хищных животных несомненно существовавших в то время. Они появились в Казахстане, по-видимому, значительно раньше, возможно, в начале четвертичного периода или даже в плиоцене, но палеонтологических остатков их того времени в Казахстане пока не встречено. В осадках рассматриваемой эпохи были обнаружены костные остатки следующих хищников: волка Canis lupus Linnaeus, 1758; лисицы Vulpes vulpes Linnaeus; пещерного медведя Ursus (Spelaearctos) rossicus Borissiak, 1930; бурого медведя Ursus (Ursus) arctos Linnaeus, 1823; пещерной гиены Crocuta (Crocuta) spelaea Goldfuss, 1823 [Crocuta spelaea Goldfuss],
Волк, начиная с раннего плейстоцена, был широко распространен в Евразии (кроме крайнего юга) и Северной Америки. Этот эвризональныйу или повсеместный, вид обитал повсюду от тундр до пустынь. На территории Казахстана его остатки пока найдены на западе и северо-востоке. Вместе с ним в восточной части нашей республики обитали бурый медведь, барсук, горностай, ласка и др.
{kind=link}
Остатки обыкновенной лисицы найдены в пещере Уш-бас Джамбулской области и по низовью р. Урала. Лисица предпочитает открытые ландшафты равнин и горных районов. Пещерная гиена (рис. 33) Европы и Северной Азии — близкая родственница пятнистой гиены Африки. По черепам они бывают неотличимы, разве что у пещерной все кости массивнее, череп и зубы крупнее. Вся сила гиен в челюстях, массивных дробящих зубах и шейных мускулах. Это типичные трупоеды, способные разгрызать кости почти всех крупных зверей. Пещерная гиена жила в открытых скалистых местностях, поросших травой и кустарником, и питалась падалью, обычно трупами копытных (Верещагин, 1979). Ее костные остатки находят в пещерах Восточного Казахстана. Большинство погрызов на костях слонов и копытных из слоев четвертичного периода в средней полосе Евразии сделано этими гиенами.
Малый пещерный медведь (рис. 34), как считает Н. К. Верещагин (1979), не был предком современного бурого медведя, а так сказать, «двоюродным братом»— боковой тупиковой ветвью, полностью вымершей в конце последней ледниковой эпохи.
{kind=link}
Этот медведь примерно на Уз мельче большого пещерного медведя, т. е. ростом с современного бурого средней величины и масса его достигала 120—150 кг. Он был менее подвижным и более растительноядным, чем его гигантский собрат. Окраска пещерных медведей, вероятно, была буровато-белесой или соломенно-желтой, а их жизнь в основном проходила в пещерах. Там они отдыхали после жировки, сюда же забирались на зимнюю спячку, выводили молодых, играли, здесь же они и умирали. Свирепые охотники каменного века постоянно разыскивали теплые обжитые медведями пещеры и, убивая хозяев, селились в них сами. У охотников каменного века пещерные медведи были своего рода мясным скотом.
Остатки этого вида находили в степях Предкавказья, юж него Приуралья, в предгорьях Среднего Урала, Казахстана (окр, г. Зыряновска), в степях Западной Сибири и на Алтае.
Многочисленны остатки мелких млекопитающих в верхнеплейстоценовых отложениях. В их числе Citellus (Colobotis) erythrogenys Brandt, 1841—краснощекий суслик; Citellus (aff. Colobotis) pygmaeus Pallas, 1778 — малый суслик; Citellus (Urocitellus) undulatus Pallas, 1778 — длиннохвостый (азиатский) суслик; Marmota bobac Muller, 1776 — степной сурок (байбак); Allactaga jaculus Pallas — большой тушканчик; Alactagulus pygmaeus Pallas, 1778 — тарбаганчик (земляной зайчик); Dipus sagitta Pallas, 1773 — мохноногий тушканчик; Scirtopoda telum — обыкновенный емуранчик; Ellobius (Ellobius) talpinus — обыкновенная слепушонка; Myospalax myospalax Laxmann, 1773 — алтайский цокор; Lagurus lagurus — степная пеструшка; Eolagurus luteus — желтая пеструшка; Clethrionomys rufocanus Sundervall, 1846—1847 — красно-серая полевка; Arvicola terrestris Linnaeus, 1758 — водяная полевка (водяная крыса).